Article Text

Abstract
Objectives Achilles tendinopathy (AT) is a multifactorial condition for which genetic risk factors have been identified. A pathway-based approach was used to investigate genes within the inflammatory pathway.
Methods Functional polymorphisms within IL-1β (−31T→C and −511C→T), IL-1RN (variable number tandem repeat) and IL-6 (−172G→C) were investigated for associations with AT in a South African (SA) and Australian (AUS) case–control studies. A total of 369 (161 SA and 208 AUS) asymptomatic control participants (CON) and 175 (90 SA and 85 AUS) participants with AT (TEN) were genotyped. Allele combinations were constructed using the above polymorphisms in combination with the COL5A1 BstUI RFLP.
Results Independently, no associations were observed between any of the polymorphisms tested and risk of TEN. The allele combinations of five polymorphisms were, however, found to have a highly significant relationship with AT (p=0.005), after adjusting for gender and country (SA or AUS).
Conclusions Variations within the interleukin genes and the COL5A1 BstUI CC genotype are collectively significantly associated with risk of AT. This research emphasises that a pathway-based genetic association study may be a more effective approach to capture and understand the genetic risk factors underlying the multifactorial conditions, such as AT.
Statistics from Altmetric.com
Introduction
Overuse Achilles tendinopathy (AT) is considered as a multifactorial condition caused by several extrinsic and intrinsic risk factors.1 The most recently identified intrinsic risk factor for AT is an individual's genetic background.2 Genetic association studies have investigated genes encoding (1) the α chain of type V collagen (COL5A1), (2) tenascin C (TNC) which are both structural matrix proteins of tendons and (3) enzymes that breakdown the matrix such as matrix metalloproteinases (MMP-3). Genes encoding proteins involved in other biological pathways related to tendon pathology such as the inflammatory process can, however, also be explored for associations with AT.3 Expression studies have shown altered expression of several inflammatory genes/proteins in tendinopathic tendons.4 A sequence variant within the gene encoding the inflammatory cytokine, growth differentiating factor 5, was associated with risk of developing AT.5 Cytokines, such as the interleukins, are major contributors to the inflammatory response and are upregulated in tendinopathy and mechanically loaded tenocytes.6 For this reason, the inflammatory genes IL-1β, IL-1RN and IL-6 were chosen as candidate genes for associations with AT.
IL-1β encodes the interleukin-1β (IL-1β) protein which, in tenocytes, causes the upregulation of cyclooxygenase-2 and prostaglandin E2.6 7 IL-1β upregulates its own expression and the expression of other cytokines such as IL-6.6 The inflammatory mediators, induced by IL-1β, upregulate the expression of proteins involved in degradation of the tendon extracellular matrix (ECM), such as the matrix metalloproteinases MMP-1, MMP-3 and MMP-13 (figure 1).6,–,8 One of the components of the ECM targeted for degradation by these MMPs is type V collagen.9 In addition, IL-1β may also affect the expression of the COL5A1 gene via transforming growth factor β (figure 1).10 11 Although these interactions have only been observed in chondrocytes and osteoblasts,10 11 it is feasible that similar mechanisms exist in tenocytes.

Proposed pathways through which increased mechanical loading induces expression of IL-1β, IL-1ra and IL-6, which in turn may act to influence the expression of type V collagen thereby potentially modulating the risk of AT.7 9,–,11 18 43 IL-1β increases MMP expression by increasing expression of inflammatory mediators such as COX-2 and PGE2.6 7 IL-6 induces tenocyte apoptosis causing the production of reactive oxygen species and subsequent activation of caspases 8 and 3.26 Some of these interactions have been observed in chondrocytes and are assumed to be similar in tenocytes. Up and down arrows refer to increased or decreased expression, respectively. Boxed molecules are those investigated in this study. The functional alleles are given in brackets and have previously been shown to affect the expression of the proteins as indicated by the arrows. IL-6, interleukin-6; IL-1β, interleukin-1β; IL-1ra, interleukin-1 receptor antagonist; MMP, matrix metalloproteinase; TGF-β, transforming growth factor β.
Two single-nucleotide polymorphisms (SNPs), −31T→C (rs1143627) and −511C→T (rs16944) within IL-1β, were associated with several multifactorial conditions12,–,14 including keratoconus, a chronic condition affecting the cornea in which mechanical injury has also been implicated to trigger ECM degradation and apoptosis.14 More specifically, the TT genotype of SNP −31T→C and the CC genotype of SNP −511C→T were associated with increased risk in these conditions.12,–,14 The IL-1β −31 T allele and IL-1β −511 C allele have been implicated in increased expression of IL-1β mRNA.15 Increased IL-1β expression may result in increased ECM degradation, that is, characteristic of AT (figure 1). IL-1β, therefore, has the potential to indirectly affect the degradation and expression of type V collagen (figure 1). The CC genotype of the COL5A1 BstUI T→C RFLP was previously associated with reduced risk of AT in two populations (South Africa and Australia).16 17 Interactions between this genotype and SNPs within the IL-1β gene may therefore collectively modulate the risk of developing AT.
The receptor antagonist of IL-1β, interleukin-1ra (IL-1ra), is encoded by IL-1RN, located downstream of the IL-1β gene on chromosome 2q14. IL-1ra competitively binds to the same IL-1 receptor as IL-1β, without inducing any effects.18 It is the ratio of IL-1β/IL-1ra that determines the biological effect of IL-1β. In this way, the levels of IL-1ra may potentially also influence tendon ECM degradation, including the degradation of type V collagen. The two-repeat allele of the variable number tandem repeat (VNTR) polymorphism (rs2234663) within the IL-1RN gene was previously associated with negative outcomes in a variety of gastrointestinal diseases,19 osteoporotic fractures18 and atherosclerosis.20 The VNTR has also been shown to be in linkage disequilibrium with IL-1β −511C→T in a single study investigating renal failure in individuals from India.21 Studies have provided conflicting evidence concerning the effect of the two-repeat allele on IL-1ra expression.22,–,25 Some of these studies report that the two-repeat allele is associated with an increase expression of IL-1ra (twofold increase) whereas others have shown the inverse relationship. Nonetheless, IL-1ra modulates the effects of IL-1β (figure 1) and an alteration in IL-1ra expression may indirectly alter the expression of type V collagen.
Interleukin-6 (IL-6) encoded by IL-6 is thought to play a role in apoptosis (figure 1).26 Tenocytes are responsible for maintenance of the ECM1 and therefore if the number of tenocytes present in the tendon is reduced, both the maintenance of the ECM of the tendon as well as its repair will be compromised. Tenocyte apoptosis, characteristic of tendinopathy, was found to be accompanied by increased expression of IL-6, as well as increased levels of the apoptotic mediators, caspase-3 and caspase-8.26 In addition, IL-6 indirectly affects the expression of the COL5A1 gene by means of a similar mechanism to that of IL-1β (figure 1).11
A functional −172G→C SNP within IL-6 was previously associated with a variety of conditions.27,–,29 Expression analysis revealed that IL-6 −172G→C results in differential transcription rates, with the G allele causing increased expression of IL-6.28 This increased IL-6 expression may lead to increased tenocyte apoptosis and has the biological potential to increase an individual's risk of developing AT. Furthermore, alterations in IL-6 expression may indirectly potentially also regulate type V collagen expression (figure 1).
Due to the clinical and functional associations described above, the IL-1β −31T→C and −511C→T, IL-1RN VNTR and IL-6 −172G→C polymorphisms were investigated for associations with AT in South African (SA) and Australian (AUS) individuals. Furthermore, gene–gene interactions were explored to determine how they may modify risk of developing AT.
Methods
Participants
A case–control genetic association study was conducted following a candidate gene approach. A total of 251 physically active unrelated Caucasian SA participants comprising of 161 asymptomatic control participants (SA CON) and 90 with diagnosed AT (SA TEN) were recruited for the study as previously described.16 30 31 The 293 Caucasian AUS participants comprised of 208 AUS CON and 85 AUS TEN participants and were recruited as previously described.17 All the participants signed the informed consent forms according to the Declaration of Helsinki, provided personal particulars and completed a questionnaire regarding medical history.31 The SA and AUS participants were of self-reported European Caucasian ancestry. The profile of the CON and TEN participants of both the SA and AUS populations have been included in the supplementary information (supplement A).
Approval for the study was obtained from the Research Ethics Committee of the Faculty of Health Sciences within the University of Cape Town (reference number 172/2005) and Human Ethics Committee of La Trobe and Deakin Universities, Melbourne, Australia.
DNA extraction
DNA was extracted for the SA participants30 and the AUS participants17 as previously described.
Polymorphism analysis
Three genes, IL-1β, IL-6 and IL-1RN, were chosen for analysis because of their role in the inflammatory pathway and their biological relevance to tendon pathology.6,–,8 26 32,–,34 Common clinically associated polymorphisms within these genes were annotated (figure 2). Three SNPs were selected for investigation; −31T→C and −511C→T are located within the promoter region of IL-1β12 13 35 and −172G→C within the promoter region of IL-6.27,–,29 36 The VNTR within intron 2 of IL-1RN was also investigated (figure 2).18,–,20 37
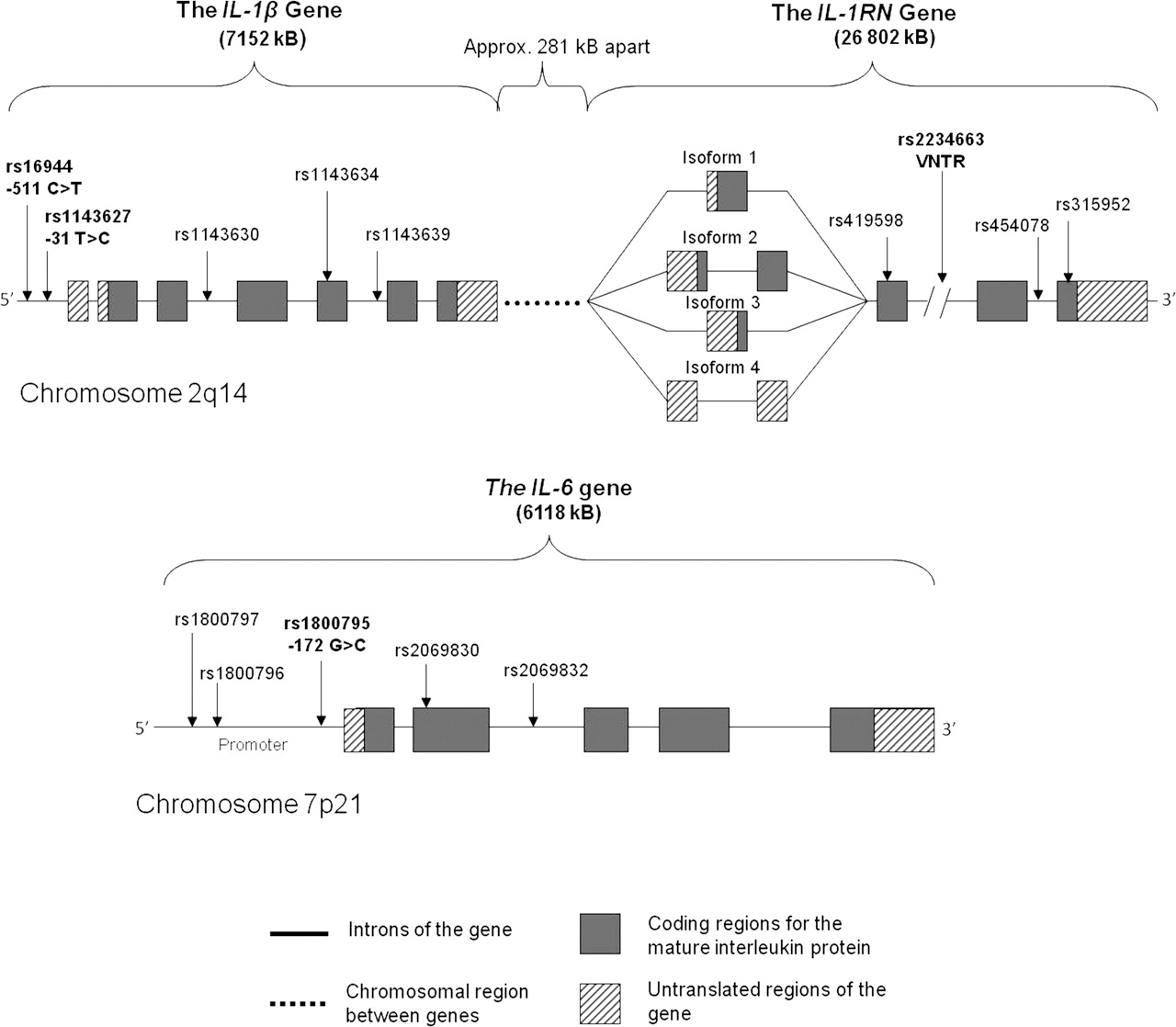
A schematic representation of the IL-1β, IL-1RN and IL-6 genes depicting some of the common clinically associated polymorphisms. Sizes of the genes are given in brackets. Major and minor alleles of the single-nucleotide polymorphisms (SNPs) are indicated in parentheses, respectively. The polymorphisms investigated in this study are in bold. Isoform 1 of the IL-1RN gene encodes the secreted form of IL-1ra and isoforms 2–4 encode intracellular forms of IL-1ra. Polymorphisms were identified and figure was constructed using information from databases hosted by the National Centre for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov/) and the Ensembl Genome Data Centre (http://www.ensembl.org/index.html).
Genotyping of IL-1β −31T→C (rs1143627), IL-1β −511C→T (rs16944) and IL-6 −172G→C (rs1800795) was conducted using PCR and restriction fragment length polymorphism (RFLP) analysis whereas the VNTR was genotyped using PCR. Polymorphism analysis was carried out following the same protocol for both SA and AUS samples. The localisation of the various SNPs, primer sequences and RFLPs were annotated (table 1 and figure B1 in supplement B).
The primer pairs used for the amplification of IL-1β −31T→C, IL-1β −511C→T, IL-6 −172G→C and the IL-1RN VNTR polymorphisms and the specific allele sizes for each polymorphism
Genotyping
The IL-1β gene
PCR amplification reactions for −31T→C and −511C→T were performed in 40 μl containing 200 ng of genomic DNA, 20 pmol of each primer (table 1), 2.0 mM MgCl2, 50 mM KCl, 10 mM Tris–HCl (pH 8.3), 200 μM dATP, dCTP, dGTP, and dTTP and 0.5 U Taq DNA polymerase (New England Biolabs, Ipswich, Massachusetts, USA). PCR was conducted using a thermal cycler (Hybaid; PCR Express, Middlesex, UK). PCR conditions were an initial denaturing step of 94°C for 10 min, 35 cycles of 94°C for 25 s, 54°C for 45 s and 72°C for 30 s and final incubation at 72°C for 5 min (adapted from Yang et al13 and Sakuma et al12). AvaI and AluI RFLPs (figure B1 in supplement B) were used for allelic discrimination of −31T→C and −511C→T, respectively, following manufacturer's recommendations (New England Biolabs). Products were resolved on 2% agarose gels and visualised by SYBR Gold staining (figure B2 in supplement B).
The IL-6 gene
IL-6 −172G→C was PCR amplified, using primers specified in table 1, following standard conditions with minor changes: SuperTherm Taq (Southern Cross Biotechnology, Claremont, South Africa) and 3.5 mM MgCl2 was used. Alleles were discriminated using the NlaIII RFLP (figures B1 and B2 in supplement B) (New England Biolabs).
The IL-1RN gene
PCR amplification reactions of the VNTR within intron 2 of IL-1RN (figure B1 in supplement B) were performed, using primers specified in table 1, following standard conditions. PCR cycling parameters were a denaturing step of 94°C for 5 min, 35 cycles of 94°C for 30 s, 51°C for 30 s and 72°C for 40 s, and a final extension step of 72°C for 5 min. PCR products were resolved on 2% agarose gels (figure B2 in supplement B). Five alleles of the VNTR are presented in table 1.12 38
Statistical analysis
Basic characteristics of the study groups were presented in previous published studies16 17 30 31 and are summarised in appendix A. Logistic regression was used to compare the population groups (SA and AUS) with respect to baseline characteristics as well as genotype and allelic distributions. In order to increase power and reduce the number of individual tests done, the population groups were combined for analysis of diagnostic status and therefore all analyses were also adjusted for country (SA and AUS). Allele combinations were inferred for each possible selection of genotypes, from pairs to all five of the polymorphisms investigated in this study. Logistic regression was also used to compare the TEN and CON groups with respect to genotype, allele and allele-combination frequencies. All analyses were adjusted for the interaction between gender and population group because it was found to have a highly significant association with diagnostic group.
Results corresponding to a p value of <0.05 were described as significant. The freely available programming environment, R39 and R packages were used for all analyses. The R package genetics40 was used to estimate genotype and allele frequencies and Hardy–Weinberg equilibrium (HWE) probabilities. Frequencies of allele combinations (of polymorphisms) were inferred and analysed using the R package, haplo.stats.41 42
Results
Participant characteristics
The SA and AUS participants investigated in this paper were previously described and compared in detail (supplement A).16 17 30 31 The gender distribution differed highly significantly between the SA and AUS groups and a highly significant interaction between gender and group (SA and AUS) on diagnosis was also noted (table A2 in supplement A). The percentage of women was the same in both SA and AUS TEN groups, but differed significantly in the CON groups (p<0.001). The percentage of women in the AUS TEN group are less than half (27%) of the percentage of women in the AUS CON (60%) group, whereas in SA the difference is much smaller between the TEN and CON groups. The IL-6 −172G→C genotype and allelic distributions differed significantly between SA and AUS. No other genotype effects were observed on the descriptive measures (results not shown).
Genotype and allele frequency distributions
No significant differences in the distributions of the genotype or allele frequencies at the IL-1β −31T→C, IL-1β −511C→T and the IL-1RN VNTR loci were noted (1) between the two countries (SA and AUS) after adjusting for gender and diagnoses or (2) between diagnoses, adjusting for gender and country and (3) between genders, adjusting for diagnoses and country (SA and AUS) (table 2). A significant difference in the genotype frequency distribution (p=0.023) and the allele frequency distribution (p=0.006) was, however, noted at the IL-6 −172G→C locus between the two countries, after adjusting for gender and diagnoses (table 2). In the control group, the observed (minor) C allele frequency was 38% for SA and 49% for AUS. The estimated odds of each C allele in SA is 30% lower than in AUS; OR=0.70; 95% CI: 0.54 to 0.90. All groups were in HWE except for the AUS CON group at the IL-1β −31T→C. The frequency distributions of the individual genotypes and minor alleles for both SA and AUS (table 2) were similar to the frequencies reported for the European populations but interestingly they were significantly different between the countries for IL-6 −172G→C SNP (http://www.ncbi.nlm.nih.gov/projects/SNP/).
Genotype frequency distributions and minor allele frequencies of IL-1β −31T→C (rs1143627), −511C→T (rs16944) and IL-6 −172G→C (rs1800795) SNPs and the VNTR in control (CON) and Achilles tendinopathy (TEN) groups of South Africa (SA) and Australia (AUS)
Interactions
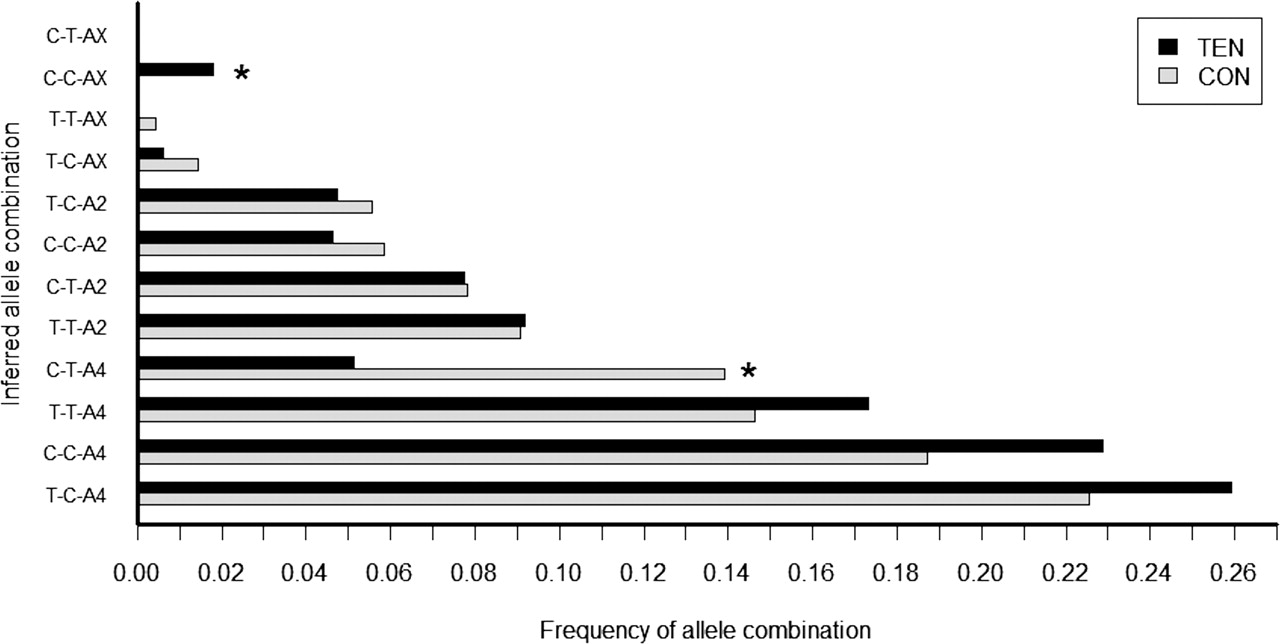
Frequencies were inferred for the allele combinations of the COL5A1 BstUI RFLP – IL-1β −31T→C – IL-1β −511C→T – IL-6 −172G→C – IL-1RN VNTR polymorphisms genotyped in the combined SA and AUS groups. The frequency distributions in TEN and CON are graphed below (figure 3). The most common inferred allele combinations for five polymorphisms genotyped in both the TEN and CON groups was T-T-C-G-A4. The allele combinations of five polymorphisms were found to have a highly significant relationship with TEN risk (p=0.005), after adjusting for gender and country (SA or AUS). More specifically, both T-T-C-G-A2 (p=0.017; 4% vs 1%) and C-T-C-G-AX (p=0.006; 2% vs 0%) had higher frequencies in TEN, after adjusting for country, gender and their interaction. Analysis was done using a score test, which provides a score statistic that cannot be directly interpreted as an effect size. The frequencies of the allele combinations are too low to estimate the adjusted ORs with any precision.

The frequency distribution of the five-way allele combinations of the COL5A1 BstUI RFLP – IL-1β −511C→T – IL-6 − 172G→C – IL-1RN VNTR polymorphisms genotyped in the combined South African (SA) and Australian (AUS) control (CON) and Achilles tendinopathy (TEN) groups. The allele combinations are ordered according to their frequencies in the control group. The rare alleles of the VNTR, A5 and A1, were combined and called AX. If the TEN bar is shorter than the CON bar, the inferred allele combination might be ‘protective’ and if longer it might be a ‘risk’ haplotype. The asterisks denote the statistically significant allele combinations associated with the TEN (C-T-C-GA-X and T-T-C-G-A2) group.
The frequencies of the inferred allele combinations of all the smaller number of polymorphisms (2, 3 and 4) were examined and only those with significant global p values are reported. The four-way combination (COL5A1 BstUI RFLP – IL-1β −511C→T – IL-6 −172G→C – IL-1RN VNTR) was significantly associated with TEN risk (p=0.014) after adjusting for gender and country (SA or AUS). More specifically, C-T-C-A4 (p=0.045; 9% vs 3%) was more prevalent in CON than TEN (figure 4). In contrast, T-C-G-A2 (p=0.031; 4% vs 1%) and C-C-G-AX (p=0.017; 2% vs 0%) were more prevalent in TEN than CON.

The frequency distribution of the four-way allele combinations of the COL5A1 BstUI RFLP – IL-1β −511C→T – IL-6 − 172G→C – IL-1RN VNTR polymorphisms genotyped in the combined South African (SA) and Australian (AUS) control (CON) and Achilles tendinopathy (TEN) groups. The allele combinations are ordered according to their frequencies in the control group. The rare alleles of the VNTR, A5 and A1, were combined and called AX. If the TEN bar is shorter than the CON bar, the inferred allele combination might be ‘protective’ and if longer it might be a ‘risk’ haplotype. The asterisks denote the statistically significant allele combinations associated with either the TEN (C-C-G-AX and T-C-G-A2) or CON (C-T-C-A4) group.
The three-way combination (COL5A1 BstUI RFLP – IL-1β −511C→T – IL-1RN VNTR) also had a significant (p=0.040) relationship with TEN, after adjusting for gender, country and their interaction. The C-T-A4 inferred allele combination was more prevalent in CON than TEN (p=0.013; 14% vs 5%) whereas the C-C-AX was less prevalent (p=0.026; 0% vs 2%) (figure 5). The odds of TEN with the C-T-A4 combination is 61% less than with ‘wild type/most frequent’ T-C-A4 (OR=0.39; 95% CI: 0.18 to 0.83).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The frequency distribution of the three-way allele combinations of the COL5A1 BstUI RFLP – IL-1β −511C→T – IL-1RN VNTR polymorphisms genotyped in the combined South African (SA) and Australian (AUS) control (CON) and Achilles tendinopathy (TEN) groups. The allele combinations are ordered according to their frequencies in the control group. The rare alleles of the VNTR, A5 and A1, were combined and called AX. If the TEN bar is shorter than the CON bar, the inferred allele combination might be ‘protective’ and if longer it might be a ‘risk’ haplotype. The asterisks denote the statistically significant allele combinations associated with either the TEN (C-C-AX) or CON (C-T-A4) group.
Discussion
This study followed a pathway-based approach to investigate the association of sequence variants within IL-1β, IL-1RN and IL-6 and AT. The most interesting finding was the significant interactions between sequence variants within these genes and the COL5A1 BstUI T→C RFLP which, in combination, are significantly associated with risk of developing AT. This observation provides preliminary evidence that genetic factors potentially influence the inflammatory pathway in the pathogenesis of AT.
The body of evidence for the involvement of cytokines such as IL-1β and IL-6 and other inflammatory mediators in the development of tendinopathy is growing.4 Studies have demonstrated that inflammatory gene expression profiles of tenocytes are modulated in response to mechanical loading26 and that these cytokines trigger tenocyte apoptosis and pathological ECM degradation.7 8 26
The current study did not find an association with the functional variants within the IL-1β,IL-6 or IL-1RN genes and symptoms of AT in either of the population groups. However, the inferred allele combinations constructed from the COL5A1 BstUI RFLP – IL-1β −31T→C – IL-1β −511C→T – IL-6 −172G→C – IL-1RN VNTR polymorphism analysed in the combined SA and AUS groups suggest the potential interactions of these respective sequence variants in modulating the risk profile of developing AT.
The exact interactions between the COL5A1 gene and these interleukin genes are not yet described. However, it is interesting to note that type V collagen may be regulated by certain inflammatory mediator proteins in the IL-1β-signalling pathway (figure 1).43 The upregulation of COL5A1 gene expression could potentially result in an increase of α1(V) collagen chain, which may cause decreased tendon fibril diameter.44 This modulation of expression together with the increased IL-1β-mediated degradation of type V collagen could hypothetically result in weaker tendons and an increased risk of AT.
To determine the true population effect size of these variants on the risk of developing AT, large prospective longitudinal studies need to be conducted. Investigating the allele combinations at these five loci needs further exploration in other independent populations and if repeated could potentially highlight the significance of these functional gene polymorphisms in the development of the condition and thereby point to the biological pathways, such as the inflammatory pathway, which may be involved in the aetiology of AT. More importantly, further proof of the association of these genotype profiles may highlight the potential of these loci to be used as biomarkers in a risk model for tendinopathy.
Key components of these pathways could potentially be targeted for biology-based therapeutic intervention strategies to prevent/reduce the progression of AT. We are in the era of personalised medicine and clinicians are already making use of an individual's unique genetic architecture to tailor treatment and identify risk for disease.45 Some of the therapies which could be administered include the local administration of anticytokine antibodies, soluble cytokine receptors or cytokine antagonists.26 All three strategies would aim to decrease the biological effect of the cytokine and thus reduce the ECM degradation, preventing pathological tendon degeneration.
One limitation to this study was that the TEN groups were significantly heavier than the CON groups at recruitment. This difference may, in part, be due to the age difference between the TEN and CON groups, as the TEN groups were older than the controls at the time of recruitment. Accurate weight measurements for the SA and AUS TEN groups at the time of the injury could not be obtained, however, a number of individuals in the TEN groups anecdotally reported increased weight and decreased physical activity after development of AT. Increased adiposity has been reported in many studies of tendinopathy46 and has also increased central fat in men with AT pathology who had no history of pain suggests that increased adiposity may be a risk factor for tendon injury.47 It is possible that the tendinopathy is mediated through increased inflammatory cytokines associated with increased adiposity. Also, there were significantly more males in the AUS TEN group. In accordance, after adjusting for participant age at recruitment or gender, the difference in weight between the CON and TEN groups was no longer significant.
In conclusion, our novel findings suggest that interactions between interleukin genes and the COL5A1 BstUI CC genotype polymorphism may potentially modulate the risk of AT. This study highlights the need for further exploration of the inflammatory pathway in the attempt to understand the biological mechanisms underlying the pathophysiology of AT. It also emphasises that a pathway-based genetic association study is more effective in capturing and understanding the genetic risk factors underlying a complex phenotype compared to the single candidate gene approach. Furthermore, this study provides evidence that injury susceptibility is most likely dependent on the cumulative effect of the interactions of sequence variants in multiple genes within biologically significant pathways.
What is already known on this topic
Sequence variants within four genes have been associated with Achilles tendinopathy. Achilles tendinopathy is a degenerative condition, however, there is a growing body of evidence suggesting the involvement of regulators of the inflammatory pathway to be altered in tendinopathic samples.
What this study adds
This association study provides preliminary evidence that the inflammatory pathway is possibly one of the significant biological pathways involved the pathogenesis of Achilles tendinopathy and requires further exploration.
Acknowledgments
The authors would like to thank Dr GG Mokone for assistance in the recruitment of the SA control and Achilles tendinopathy participants.
References
Supplementary materials
Web Only Data bjsm.2010.076760
Files in this Data Supplement:
Footnotes
EN and KO'C contributed equally to this work
-
Funding This study was supported in part by funds from the National Research Foundation (grant numbers SUR2008060500012 and FA2005021700015), the South African Medical Research Council, the University of Cape Town and Discovery Health. The final preparation of this study for publication was supported in part by the International Olympic Committee (IOC) Research grant to the Clinical Sports Medicine Group of the UCT/MRC Research Unit for Exercise Science and Sports Medicine of the University of Cape Town. The authors, EN and KO'C, were awarded the grantholder-linked student support from the National Research Foundation (NRF) of South Africa (Grant SUR2008060500012).
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval This study was conducted with the approval of the University of Cape Town, and the University of La Trobe and Deakin University
-
Provenance and peer review Not commissioned; externally peer reviewed.